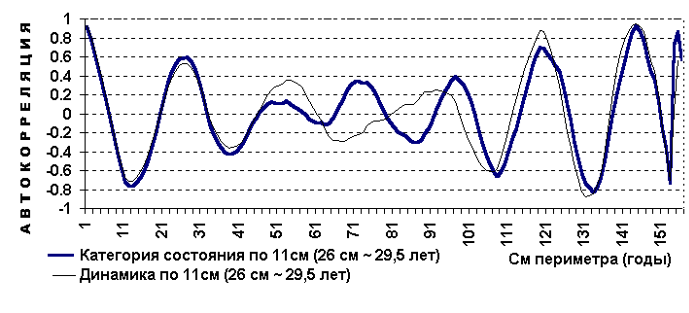
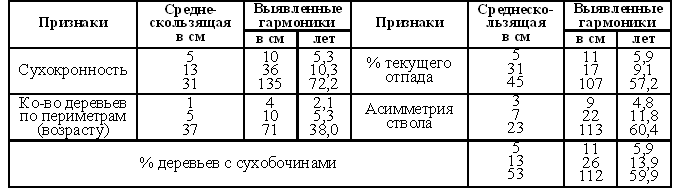
................................................................................................................................................................................................................ 2. Тема. Периодичность фенотипа на уровне популяции. Чижевский А.Л. [17], отметил периодичность микроорганизмов. Выявлена периодичность усыхания насаждений [18, 19]. Отмечена периодичность численности насекомых, зайца и рыси. Маслов С.Ю. [20] утверждал, что доминирование левого или правого полушария мозга в популяциях человека различно в различные исторические эпохи. Гумилев Л.Н. [21] писал, об этом же явлении, но о гармониках с большим периодом. Этим же явлением объясняется смена реалистичных наскальных рисунков периода палеолита схематично абстрактными рисунками в неолите (еще большая гармоника периодичности). Демограф Зигфрид Коллер [22] утверждал, что есть более жизнеспособные когорты поколений и менее жизнеспособные. Четвериков С.С. [23] определил подобные явления как «волны жизни». Все выше перечисленные периодичные признаки полигенны и априори задаются периодичными полигенными физиологическими процессами и периодичной активностью генов неких комплексов генов. Автор обнаружил подобные же периодические явления полигенных признаков в популяциях древесных пород и у индивидуума – мужчины и в таксонах большего порядка чем вид, что отражено в 3 теме. .В 1981 году при лесопатологическом обследовании Тульских засек, на множестве пробных площадей отмечена синхронная альтернация сухокронности дуба (Таблица 7, 8) по когортам поколений. Сухокронность – признак реакции дерева на отрицательные факторы среды и определяется категорией или классами состояния по проценту сухих ветвей в кроне. Диаметр или кумулятивный радиальный прирост дерева подчиняется логистической кривой [24, 25]. В средней части эта функция почти прямолинейна и прирост равен определенной единице времени. Таблица 7. Альтернация сухокронности дуба. Яснополянское лесничество квартал 23 выдел 21. 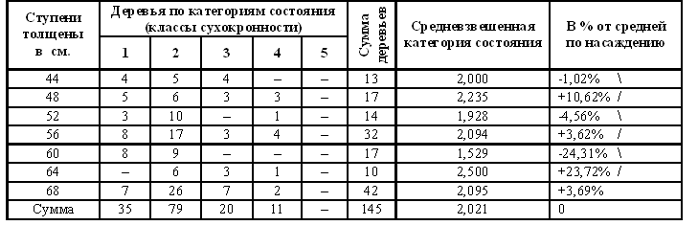 На постоянной пробной площади (Таблица 8) отмечена аналогичная альтернация, но здесь представлена только ее динамика между 1980 - 1981 гг. Если категория сухокронности между учетами не изменилась, ставился балл 0, если происходило ухудшение, ставился балл равный изменению категории со знаком минус, если улучшение – то плюс соответствующий балл. Таблица 8. Альтернация динамики сухокронности дуба. Музей «Ясная поляна», Арковский верх. 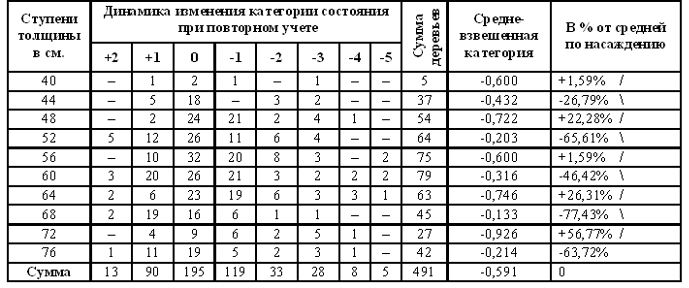 Синхронность альтернации категорий сухокронности на множестве проб подразумевает существование периодического явления во всей популяции. Категория сухокронности в 1981 году хуже у деревьев 40 – 48 – 56 – 64 – 72 ступеней толщины и лучше у 44 – 52 – 60 – 68 – 76 ступеней толщины. Ступень толщины это градация времени, случайно совпадающая на момент измерения с определенными возрастными когортами деревьев. Цикл по данному признаку равен 4 см + 4 см = 8 см. Средний годичный радиальный прирост дуба по спилам в среднем равен 2,8 мм, что согласуется с таблицами хода роста [25]. Соответственно гармоника в реальном времени примерно равна 8см / 2,8мм ≈ 29 годам, а полупериод равен 14,5 годам (4 см). В 2006 г. при обследовании Калининградской области у дуба был выявлен более точно полупериод в 4,1 см диаметра, так как у деревьев измерялся периметр на высоте груди. Далее для обнаружения гармоник периодичности применялся автокорреляционный анализ. Если ряд значений прерывался отсутствием данных, то ставилось среднее значение между двумя ближайшими. Ряд значений преобразовался в кумулятиву, тренд которой описывался функцией. Затем эта функция вычленялась, образуя некумулятивные остатки для устранения возрастного тренда. По остаткам и их усредненным рядам нечетными средне скользящими (3, 5 и т.д.) производился автокорреляцией анализ с отбором наиболее регулярных кривых. При этом сглаживаются случайные флуктуации и гармоники, меньше и равные среднескользящей, выявляя большие гармоники. Чем больше средне скользящая осреднения, тем меньше проявление "Белого статистического шума" [26], зависящего от количества элементов в группах выборки. Группами выборки в этом случае являются средне скользящие, состоящие из множества элементов выборки. Эта методика опробована на периодичности солнечной активности и показала свою эффективность при обнаружении уже известных гармоник периодичности. В 1993 году в Тульских засеках была заложена сеть постоянных пунктов учета лесопатологического мониторинга . Повторные учеты проводились в 1994 и 1997 годах. Так как явление характерно для популяции в целом, данные объединенялись в единый массив по породам. Измерялся периметр дерева на высоте 1,3 м. Проанализировано 751 дерево дуба. Среднегодичный прирост деревьев второго бонитета равен 2,8 мм [25] или 8,8 мм периметра. Таблица 9. Периодичность дуба по полигенным признакам фенотипа. 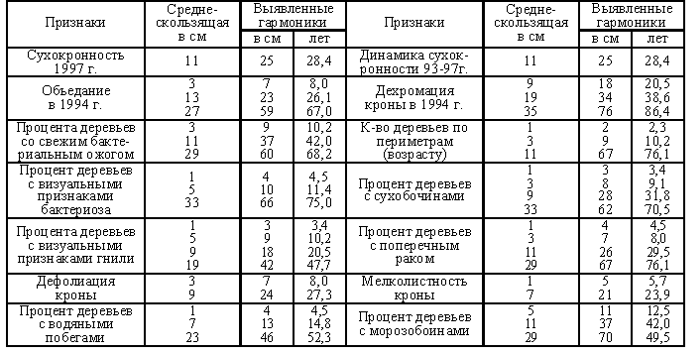 Рис. 14. Дуб. Автокорреляция категории состояния кроны и ее динамики.
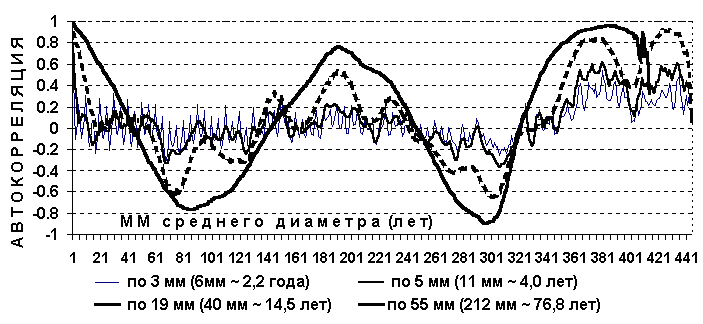
На Рис. 14 при средне скользящей в 11 см периметра обнаруживается гармоника в 26 см периметра (8,3 см диаметра), что соответствует 29,5 годам по таблицам хода роста, а полупериод 14,8 годам или 13 см периметра (4,1 см диаметра). Эта периодичность и была независимо выявлена в 1981 году на территории музея "Ясная поляна" и в Тульских засеках Таблица 7, 8 и в Калининградской области в 2006 г. более точно подтверждена.
Рис. 16. Автокорреляция процента деревьев дуба. со свежим бактериальным ожогом ствола [27, 28].
Рис. 17. Автокорреляция процента мелколистных деревьев дуба.
Мелколистность – признак феноформ ранневесеннего распускания [29]. В первую очередь объедаются определенные когорты поколений. На этих же постоянных пунктах учета проанализировано 1304 дерева липы. В таблицах хода роста по липе данных нет. Таблица 10. Периодичность липы по полигенным признакам фенотипа. 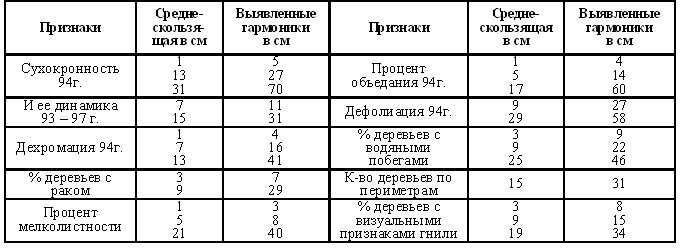 Рис. 18. Автокорреляция количества деревьев липы по периметрам (возрасту). Рис. 18. Автокорреляция количества деревьев липы по периметрам (возрасту).
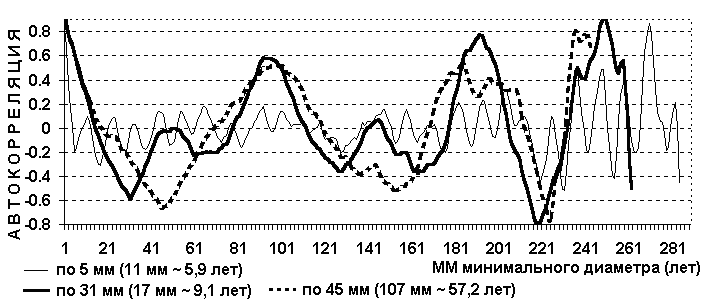
В Бурятской АССР в 1984 году в очаге сибирского шелкопряда заложена пробная площадь, на которой проанализировано 1885 деревьев кедра. Измерялся минимальный диаметр (мм) и периметр дерева (см) на высоте 1,3 м. Среднегодовой прирост четвертого бонитета равен 2,0 мм [25]. Минимальный диаметр меньше среднего на 6,31 %, и соответственно среднегодовой прирост по минимальному диаметру равен 1,87 мм. Таблица 11. Периодичность кедра по полигенным признакам фенотипа.
Рис. 19. Автокорреляция процента текущего отпада (смертность) кедра.
Текущий отпад – усыхающие деревья и свежий сухостой текущего года. Сухокроность и гибель дерева – два разных полигенных признака, так как у них не совпадают гармоники периодичности, тоже наблюдается и по пихте. Асимметрия ствола – это отношение среднего к минимальному диаметру. На этой же пробе проанализировано 886 деревьев пихты. В таблицах хода роста данных по пихте нет. Таблица 12. Периодичность пихты по полигенным признакам фенотипа.
Рис. 20. Автокорреляция количества деревьев пихты по периметрам.
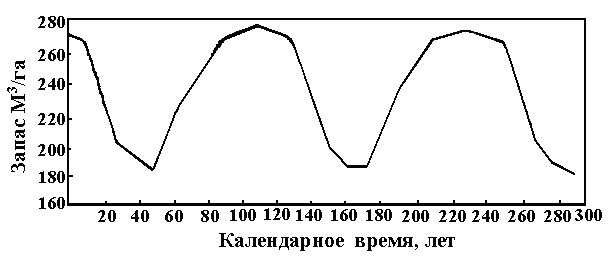
Фалалеев Э.Н. [30] отметил, что в процессе роста пихтарников запас древесины на 1 га периодичен (Рис. 21) и это отражает относительное количество деревьев по когортам поколений, что в сущности и соответствует максимальной гармонике Рис. 20. Рис. 21. Периодичность древесного запаса пихтарников Западно-Сибирской равнины [30].
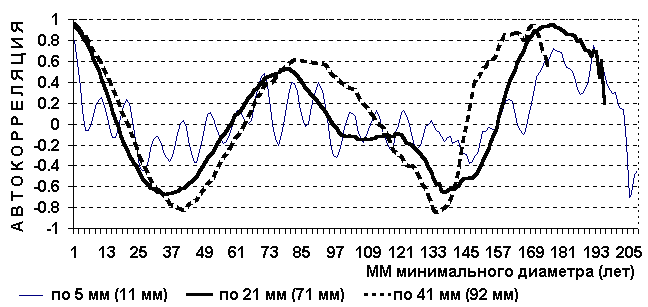
В 1991 году в Рузском лесхозе Московской области заложено 8 пробных площадей. Проанализировано 2616 деревьев ели. Измерялся периметр дерева на высоте 1,3 м и затем преобразовывался в диаметр. Средний годичный прирост деревьев второго бонитета равен 2,76 мм [25]. Таблица 13. Периодичность ели по полигенным признакам фенотипа.
Рис. 22. Автокорреляция количества деревьев ели по диаметру.
Первые две гармоники на Рис. 22 в сущности отражаю обычные периоды плодоношения ели и незначительная ошибка в 0,2 года по всей вероятности вызвана несоответствием прироста и грубой оценкой понятия бонитет. Наибольшая гармоника отражает периоды популяционного угнетения данной породы, в это время в насаждение вклиниваются породы с меньшими периодами периодичности соответствующего угнетения. В лесоводстве это явление называется сменой пород (Ель сменяется Березой, Дуб - Липой и т.п.) [31]. Рис. 23. Автокорреляция относительной протяженности кроны деревьев ели.
Объем кроны априори, пропорционален объемам физиологических процессов дерева и скорости метаболизма. У человека это отражает работа печени. Выявлено, что скорость работы печени у людей в популяции имеет двуглавую зависимость [32, 33]. По всей вероятности, это вызвано необъективным объединением в выборку четных и нечетных когорт поколений, имеющих разные генеральные совокупности этих норм. Периодичностью норм полигенного признака объясняется не детерминация повторных опытов в биологии. Например, при поиске вещества памяти [40, 41]. При учете когорт поколений решается проблема контроля и детерминации результатов различных исследований в биологии. Для анализа прочих параметров периодичности должен применяться цифровой спектральный анализ [34], но наши данные заведомо искажают амплитуду периодичности систематическими и случайными погрешностями прироста. Поэтому данную работу надо рассматривать, как предварительную и эти явления следует проанализировать по реальному возрасту деревьев на вырубках или по картотекам поликлиник для человека. Все проанализированные полигенные признаки периодичны и можно по всей вероятности предпологать, что любые полигенные признаки так или иначе периодичны, включая и у индивидуума. Периодичность визуально очень ярко прослеживается у дуба по состоянию кроны и ее динамики на простых графиках. На прочих графиках это, как правило, менее заметно и если визуально периодичность просматриваются то как правило только на больших и средних гармониках периодичности. К примеру, на графике солнечной активности также не различимы визуально малые гармоники периодичности меньше 11 летней. В популяциях исключением является периодичность по количеству деревьев ели, где и малые гармоники визуально заметны, только трудно отличить в череде циклов регулярность двух годовых от 4 годовых. По всем проанализированным породам и их полигенным признакам выявлены независимые периодичности, состоящие, как правило, из множества гармоник. Соответственно, не вдаваясь в химико-генетическую сущность, можно построить карты периодичности норм полигенных признаков популяций и изучать их «часовой» механизм эмпирически на химико-генетическом уровне. Периодичность фенотипа априори зависит от периодичной активности генов. Это контролирует: ген-оператор, ген-репрессор, ген-модификатор, ген-мутатор [35]. Системно эти гены или подобные механизмы могут периодически менять активность генофонда индивидуума, популяции, рода или всей живой материи, подчиняться периодическим "часовым механизмам" ДНК [36, 37, 38]. Последовательности оперонной регуляции для каждого полигенного признака разные соответственно разные гармоники периодичности. Вероятно в этой теме я обнаружил явления и конкретизировал их, на основании чего возможно изучение закономерностей полигенной наследственности. Для обсуждений возникших вопросов перейдите на лист обсуждений по этой теме. Обсуждение 2 темы на основании Ваших комментарий и замечаний Для личных сообщений vladimir.safronov@yandex.ru |
|
|
|